ХОРИОНИЧЕСКИЙ ГОНАДОТРОПИН
(ХГ,
хориогонадотропин), белковый гормон приматов.
ХГ - гликопротеин (мол. м. ок. 38 тыс.),
молекула к-рого состоит из двух разл. субъединиц (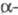 ХГ
и
ХГ
и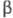 -ХГ),
нековалентно связанных друг с другом. Молекулы ХГ человека и разных видов
животных, обладая значит. гомологией, совпадают не полностью.
-ХГ),
нековалентно связанных друг с другом. Молекулы ХГ человека и разных видов
животных, обладая значит. гомологией, совпадают не полностью.
Субъединица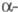 ХГ
идентична
ХГ
идентична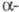 субъединице
лютеинизирующего гормона
, фолликулостимулирующего гормона
, а также
тиреотропного гормона
и включает 92 аминокислотных остатка.
субъединице
лютеинизирующего гормона
, фолликулостимулирующего гормона
, а также
тиреотропного гормона
и включает 92 аминокислотных остатка.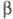 -ХГ,
полипептидная цепь к-рой состоит из 145 аминокислотных остатков, специфична
для данного гормона, но проявляет высокую степень структурной гомологии
(ок. 80%) с
-ХГ,
полипептидная цепь к-рой состоит из 145 аминокислотных остатков, специфична
для данного гормона, но проявляет высокую степень структурной гомологии
(ок. 80%) с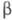 -субъединицей
лютеинизирующего гормона, отличаясь от последней удлинением С-концевого
участка полипептидной цепи на 24 аминокислотных остатка.
-субъединицей
лютеинизирующего гормона, отличаясь от последней удлинением С-концевого
участка полипептидной цепи на 24 аминокислотных остатка.
На углеводную часть, характеризующуюся
значит. гетерогенностью, приходится ок. 30% мол.м. ХГ. В ее состав входят
сиаловая к-та, L-фукоза, D-галактоза, D-манноза, N-ацетилглюкозамин и N-ацетилгалактозамин.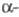 ХГ
содержит две олигосахаридные цепи, присоединенные к полипептидной цепи
с помощью N-гликозидной связи между N-ацетилглюкозамином и амидной группой
двух остатков аспарагина.
ХГ
содержит две олигосахаридные цепи, присоединенные к полипептидной цепи
с помощью N-гликозидной связи между N-ацетилглюкозамином и амидной группой
двух остатков аспарагина.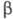 -ХГ
содержит 6 олигосахаридных цепей, 2 из к-рых присоединены N-гликозидной
связью по остаткам аспарагина, а 4 -О-гликозидной связью между остатками
N-ацетилгалактозамина и группой ОН остатков серина С-концевого участка
полипептидной цепи. Углеводные компоненты ХГ необходимы для соединения
субъединиц, поддержания конформации его молекулы, защищают полипептидные
цепи субъединиц от расщепления протеолитическими ферментами
. Удаление
углеводных остатков приводит к значит. уменьшению периода полужизни ХГ
в организме.
-ХГ
содержит 6 олигосахаридных цепей, 2 из к-рых присоединены N-гликозидной
связью по остаткам аспарагина, а 4 -О-гликозидной связью между остатками
N-ацетилгалактозамина и группой ОН остатков серина С-концевого участка
полипептидной цепи. Углеводные компоненты ХГ необходимы для соединения
субъединиц, поддержания конформации его молекулы, защищают полипептидные
цепи субъединиц от расщепления протеолитическими ферментами
. Удаление
углеводных остатков приводит к значит. уменьшению периода полужизни ХГ
в организме.
Молекула ХГ сравнительно легко диссоциирует
на субъединицы, напр. при действии мочевины или пропионовой к-ты. Изолир.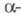 ХГ
и
ХГ
и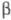 -ХГ лишены
биол. активности, но могут рекомбинировать с образованием биологически
активной молекулы ХГ.
-ХГ лишены
биол. активности, но могут рекомбинировать с образованием биологически
активной молекулы ХГ.
Специфич. биол. св-ва ХГ обусловлены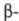 субъединицей.
Структурное сходство, имеющееся между
субъединицей.
Структурное сходство, имеющееся между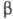 -субъединицами
ХГ и лютеинизирующего гормона, проявляется близостью их биол. и иммунологич.
св-в.
-субъединицами
ХГ и лютеинизирующего гормона, проявляется близостью их биол. и иммунологич.
св-в.
ХГ вырабатывается при беременности клетками
трофобласта (наружный слой клеток у зародышей млекопитающих) плаценты.
Может также вырабатываться нек-рыми опухолями, родственными по происхождению
клеткам трофооласта плаценты.
В процессе биосинтеза ХГ вначале на отдельных
матричных РНК синтезируется полипептидная цепь каждой из субъединиц в виде
соответствующих молекул-предшественников с более высокой мол. массой. Синтез
и присоединение олигосахаридных цепей происходят в процессе трансляции
субъединиц и после ее завершения. Зрелые полипептидные цепи ХГ образуются
путем специфич. протеолитич. модифицирования (укорочения) белков-предшественников.
Субъединицы объединяются в одну молекулу в грубом эндоплазматич. ретикулуме.
Физиол. роль ХГ состоит в поддержании
нормального развития беременности. ХГ стимулирует продуцирование желтым
телом, образующимся в яичниках при беременности, стероидных гормонов
прогестерона
и эстрогенов, что является необходимым условием для сохранения беременности.
Кроме того, ХГ участвует в регуляции др. процессов, связанных с нормальным
протеканием беременности, в т. ч. препятствует отторжению зародыша в материнском
организме, влияя на иммунные процессы. При введении ХГ в организм человека
или животных наблюдается эффект, аналогичный действию лютеинизирующего
гормона. У небеременных женщин ХГ вызывает овуляцию в созревших фолликулах
и образование желтых тел в яичниках. У мужчин ХГ стимулирует секрецию тестостерона
половыми железами.
Секреция ХГ максимальна в раннем периоде
беременности. Определение ХГ в крови или моче - основа теста на ее наличие.
ХГ выделяют из плаценты человека и животных.
Препараты ХГ для мед. применения получают из мочи беременных женщин. ХГ
используют при понижении ф-ции половых желез у мужчин и женщин, при бесплодии,
позднем половом развитии и др.
Лит.: Димитров Д. Я., Хориальный
гонадотропин человека,пер. с болт., М., 1979; Физиология эндокринной системы,
Л., 1979, с. 399; Pierce J. G., Parsons Т.F., "Ann. Rev. Biochem.", 1981,
v. 50, p. 465-95; Fiddes J.C., Talmadge K., "Recent Prog. Horm. Res.",
1984, v. 4, p. 43-78; Lustbader J. [a. o.], "J. Biol. Chem.", 1987, v.
262, № 29, p. 4204-12.
А. А. Булатов.
XOPHEPA РЕАКЦИЯ (Хорнера-Эммонса
р-ция, Уэдсворта-Эммонса р-ция, Виттига-Хорнера р-ция), получение олефинов
(в т. ч. замещенных) взаимод. альдегидов или кетонов с диалкиловыми эфирами
алкилфосфоновых к-т (диалкилфосфонатами) в присут. сильных оснований:
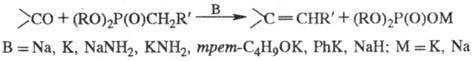
Р-цию проводят в среде орг. р-рителя (бензол,
1,2-диметоксиэтан, ДМФА, ТГФ). Обычно вначале добавляют фосфонат к суспензии
основания в р-рителе (в результате образуется фосфорилир. карбанион; см. ниже), затем вводят в р-цию карбонильное соединение. Обычно не удается
ввести в X. р. пространственно затрудненные кетоны даже при очень сильном
нагревании смеси. Выходы олефинов 60-90%. Осн. побочные продукты - смолы,
конденсир. фосфонаты.
Показано, что на первой стадии X. р. образуется
фосфорилир. карбанион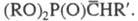 Считается, что к нему присоединяется карбонильное соед. и реализуется многостадийный
механизм, аналогичный механизму Виттига реакции
:
Считается, что к нему присоединяется карбонильное соед. и реализуется многостадийный
механизм, аналогичный механизму Виттига реакции
:
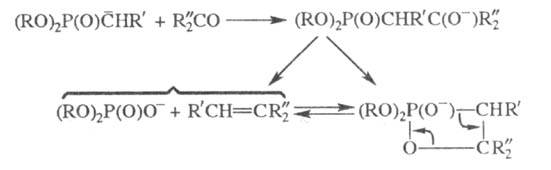
Во мн. случаях образующиеся на первой стадии
р-ции карбанионы отличаются большей реакц. способностью по отношению к
карбонильным соед., чем реактивы Виттига (фосфиналкилены).
Фосфонаты с активир. группой СН2
в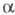 -положении
м. б. перед проведением X. р. модифицированы путем введения в это положение
атомов галогенов, алкильной группы и т. п., что позволяет синтезировать
олефины сложного строения, напр.:
-положении
м. б. перед проведением X. р. модифицированы путем введения в это положение
атомов галогенов, алкильной группы и т. п., что позволяет синтезировать
олефины сложного строения, напр.:
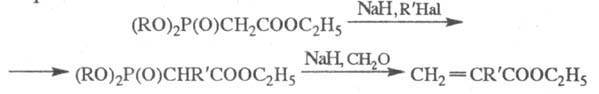
Кетены в условиях X. р. дают аллены,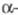 дикетоны
- сопряженные диены, оксиды олефинов - производные циклопропана.
дикетоны
- сопряженные диены, оксиды олефинов - производные циклопропана.
X. р. используют в лаб. практике. Открыта
Л. Хорнером с сотрудниками в 1958.
Лит.: Домбровский А. В., Домбровский
В. А., "Успехи химии", 1966, т. 35, в. 10, с. 1771-87; Homer L. (u. a.],
"Chem. Ber.", 1958, Bd 91, № 1, S. 61-63; Wads worth W. S., в кн.: Organic
reactions, v. 25, N. Y.-L., 1977, p. 73-253; Stec W. J., "Acc. Chem. Res.",
1983, v. 16, № 11, p. 411-17.
Г. И. Дрозд.
|


