ПРОСТАГЛАНДИНЫ
(PG),
биологически активные липи-ды, представляющие собой производные гипотетич. про-становой
к-ты (ф-ла I) и различающиеся положением заместителей и двойных связей в циклопентановом
кольце и боковых цепях.
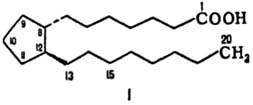
Молекулы П. имеют скелет
из 20 атомов С и содержат обычно в положении 15 гидроксигруппу. В зависимости
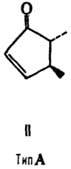
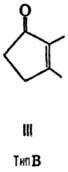
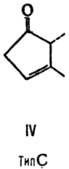
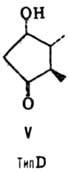
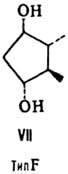
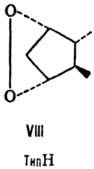
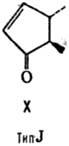
от строения цикла и характера (боковых групп в нем различают П. типов А, В,
С, D, Е, F, H, I и J (типы колец приведены на ф-лах II-X; П. G, или PGG, отличаются
от PGH наличием в положении 15 группы ООН вместо группы ОН). Цифры в ниж. индексе
букв обозначают кол-во двойных связей в боковых цепях (у П. типа F иногда в
ниж. индексе ставится также греч. буква a или b, указывающая на ориентацию
группы ОН в положении 9 относительно плоскости цикла -соотв. за или перед плоскостью
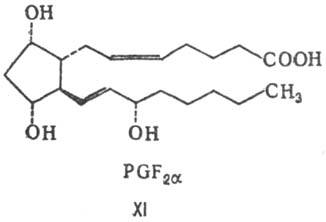
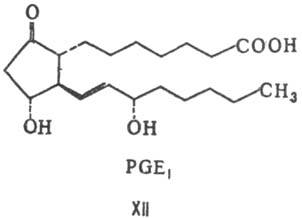
цикла)-см., напр. ф-лы соед. РGF2a (XI) и PGE1 (XII).
П. и их производные обнаружены
практически во всех клетках млекопитающих [впервые выделены из пузырьковидной
(везикулярной) железы]. Найдены также у мн. др. позвоночных и беспозвоночных
(напр., у птиц, лягушек, карпов, акул, крабов, коралловых полипов, у иек-рых
насекомых) и в ряде растений. Их содержание в большинстве тканей невелико (неск.
мкг/г и менее). Единств. богатый прир. источник П.-горгониевые кораллы (Plexaura
homo-malla), в к-рых содержание PGA2 и его производных достигает
1,5-2% от сухого веса. В кораллах найдены также биологически активные простагландиноподобные
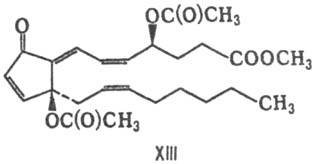
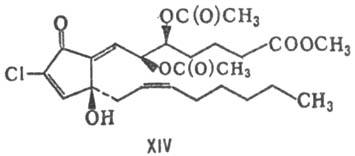
в-ва (простаноиды), отличающиеся от П. расположением функц. групп, напр. клавулон
I(ХIII) и пунагландин (XIV).

Индивидуальные П.-кристаллы
или вязкие жидкости, плохо раств. в воде, раств. в большинстве орг. р-рителей.
Для PGE1 т. пл. 115-116°С,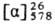 -61,6° (концентрация 0,56
г в 100 г ТГФ); для PGE2 т. пл. 66-68 °С,
-61,6° (концентрация 0,56
г в 100 г ТГФ); для PGE2 т. пл. 66-68 °С,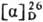 -61° (концентрация
1 г в 100 г ТГФ); для PGF2a т. пл. 30-35 °С,
-61° (концентрация
1 г в 100 г ТГФ); для PGF2a т. пл. 30-35 °С, 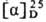 + 23,5° (концентрация 1 г в 100 г ТГФ). П., содержащие в цикле оксогруппу,
поглощают в УФ области (для П. типов А, В, С и J lмакс. соотв.
218, 278, 234 и 216 нм). Для большинства П. в кристаллич. состоянии характерна
т. наз. щпилечная конформация с приблизительно параллельным расположением боковых
цепей. П. типов Е и D легко дегидратируются в водных р-рах при рН < 4 или
рН > 8, причем в П. типа D происходит миграция транс
-двойной связи
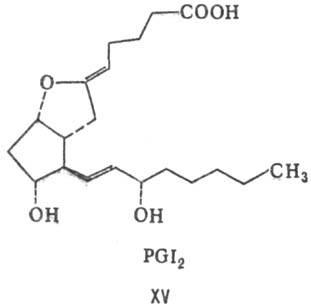
в положение 12-13. Бициклические П. нестабильны в
водных р-рах. Так, для PGI2 (простациклина, XV) период полураспада
в воде при рН 7,6 составляет 5-10 мин; он гидролизуется до 6-оксо-РGF1a.
+ 23,5° (концентрация 1 г в 100 г ТГФ). П., содержащие в цикле оксогруппу,
поглощают в УФ области (для П. типов А, В, С и J lмакс. соотв.
218, 278, 234 и 216 нм). Для большинства П. в кристаллич. состоянии характерна
т. наз. щпилечная конформация с приблизительно параллельным расположением боковых
цепей. П. типов Е и D легко дегидратируются в водных р-рах при рН < 4 или
рН > 8, причем в П. типа D происходит миграция транс
-двойной связи
в положение 12-13. Бициклические П. нестабильны в
водных р-рах. Так, для PGI2 (простациклина, XV) период полураспада
в воде при рН 7,6 составляет 5-10 мин; он гидролизуется до 6-оксо-РGF1a.
П. не накапливаются в тканях,
а синтезируются в ответ на биол. стимул из полиненасыщ. жирных к-т: П. с одной
двойной связью в боковых цепях-из эйкозатриеновой (ди-гомо-у-линоленовой), с
двумя-из эйкозатетраеновой (ара-хидоновой), с тремя-из эйкозапентаеновой (тимнодоновой)
к-т. Своб. эйкозаполиеновые к-ты (эндогенные из фосфоли-пидов или экзогенные)
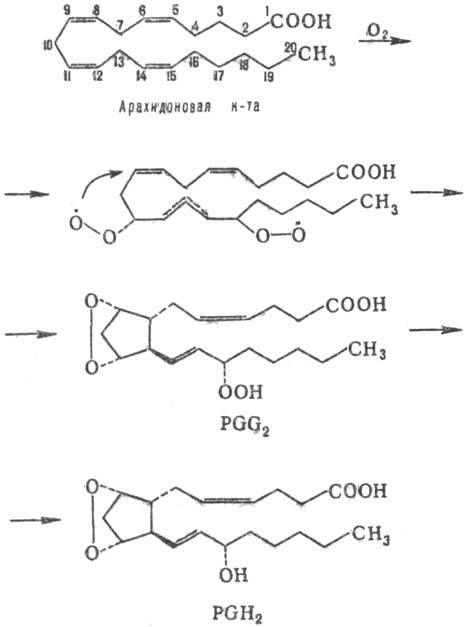
окисляются кислородом в присут. комплекса ферментов. Вначале циклооксигеназа
катализирует стереоспецифич. присоединение пероксидных радикалов в положения
11 и 15 с послед. образованием перок-сидного мостика и циклопентанового кольца;
образовавшийся PGG восстанавливается пероксидазой до более стабильного PGH -
исходного соед. для биосинтеза П. остальных типов, напр.:
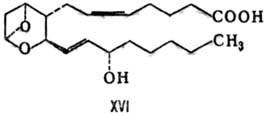
В ряде клеток из PGH2
образуется т. наз. тромбоксан (ТХ) типа А2 (ф-ла XVI)-очень нестабильное
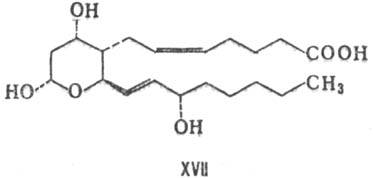
бициклич. соед. с высокой биол. активностью. В воде ТХА2 быстро гидролизуется
(период полураспада 32 с, при 37 °С) с образованием стабильного ТХВ2-ф-ла
XVII, т. пл. 95-96°С,  + 57,4° (концентрация 0,26 г в 100 г этилацетата).
+ 57,4° (концентрация 0,26 г в 100 г этилацетата).
Все П. быстро инактивируются
в организме и поэтому их концентрация в плазме кровимала (напр., 40-50 цг/мд
для PGE), тогда как
кол-во неактивных метаболитов, выделенных с мочой, может достигать 330 мкг в
сутки. Катаболизм П. начинается с их окисления НАД-зависимой 15-оксипрос-тагландиндегидрогеназой,
обнаруженной в цитозоле мн. клеток млекопитающих (наиб. активность фермент проявляет
в тканях легких, плаценты, печени, почек). Образующиеся 15-оксо-П. быстро восстанавливаются
до 13,14-ди-гидропроизводных, к-рые далее подвергаются b-
и w-окис-лению (b-окисление
- окисление карбоксилсодержащих концов боковых цепей с отщеплением от них ацетила
в виде ацетил-кофермента А, w-окисление - окисление
атома С в положении 20 до карбоксильной группы). Известны ферменты, катализирующие
взаимопревращения П. (напр., PGE2 в PGF2a,
PGA2 в PGC2).
П. обладают разнообразной
физиол. активностью, активны в низких концентрациях (10-9 М и менее).
Они участвуют в поддержании гомеостаза организма (относит. динамич. постоянства
внутр. среды и устойчивости осн. физиол. ф-ций), в воздействии на болевые рецепторы,
регулировании иммунного ответа (напр., PGE1), в родовой деятельности
(напр., PGE2 стимулирует роды, PGF2a снижает секрецию
прогестерона, необходимого для имплантации в матке оплодотворенной яйцеклетки),
поддерживают в открытом состоянии грудной проток плода во время беременности,
вызывают сокращение (П. типа F) или расширение (П. типа Е) бронхов и трахеи,
усиливают воспалит. р-цию, вызванную ожогами или др. повреждениями (способность
аспирина ослаблять воспалит. состояние связана с тем, что он необратимо ингибирует
циклооксигеназу). Кроме того, П. обусловливают повышение т-ры тела, оказывают
седатив-ное и транквилизирующее действие, стимулируют секрецию ферментов поджелудочной
железой, тормозят желудочную секрецию, способны опосредовать и модулировать
действие др. биол. стимулов, П. разных типов могут действовать как синергисты
или антагонисты. Так, баланс между уровнем простациклина PG12 (ингибирует
агрегацию тромбоцитов, расширяет артерии) и тромбоцитарным ТХА2 (индуцирует
агрегацию тромбоцитов, сужает артерии) - важный компонент гемостаза (поддерживает
постоянный состав крови). П, типов A, J и D оказывают противовирусное действие,
а П. типов J, D и их А -производные проявляют высокую прртивоопухолевую активность.
Вследствие чрезвычайно быстрого распада в организме П. действуют, в отличие
от гормонов, вблизи места секреции.
Механизмы физиол. действия
П. разнообразны. Л. взаи-мод. со специфич. рецепторами цитоплазматич. мембран,
что приводит к изменению (увеличению или уменьшению) концентрации внутриклеточных
циклич. нуклеотидов (напр., циклич. аденозинмонофосфата), способны проникать
через мембраны (включая гематоэнцефалич. барьер) и связываться С внутриклеточными
компонентами, влияя, напр., на синтез ДНК. Нек-рыс П. индуцируют перенос катионов
через биол. мембраны, изменяя физиол. состояние клеток.
Полный хим. синтез П. основан
на стереоспецифич. конденсации промежут. продуктов, содержащих фрагменты молекулы
П. Так, PGE2 с выходом 78% можно синтезировать по схеме:
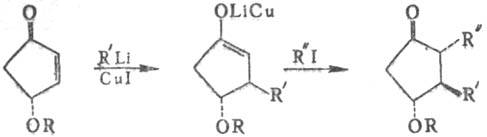
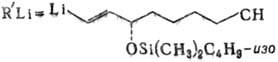

Последняя стадия в получении
PGE2-снятие защитных групп. Комбинируя промежут. продукты, получают
разнообразные аналоги П., обладающие большей стабильностью, эффективностью и
селективностью действия, чем природные.
Для количеств. определения
П., тромбоксанов и их метаболитов в биол. образцах обычно используют хроматографию
(тонкослойную, газо-жидкостную и высокоэффективную жидкостную) и масс-спектрометрию.
Наиб. точность определения достигается сочетанием методов газо-жидкост-ной или
высокоэффективной жидкостной хроматографии с масс-спектрометрией.
Препараты П. и их производных
используют в эксперим. и клинич. медицине для прерывания беременности и для
родовспоможения, терапии язвы желудка, бронхиальной астмы и нек-рых сердечно-сосудистых
заболеваний, коррекции гемостаза, как антикоагулянты при операциях с искусств.
кровообращением и при гемодиализе. Нек-рые производные П. используют для синхронизации
полового цикла при искусств. осеменении в ветеринарии.
Лит.. Варфоломеев
С. Д., Мевх А. Т., Простагландины молекулярные биорегуляторы, М., 1985; Домбровский
В. В., Грачева Е. В., Ко-чергин П. М., "Успехи химии", 1986, в.
10, с. 1720-56; Prostaglandins. Biology and chemistry of prostaglandins and
related eicosanoids, ed. P. B. Curtis-Prior, N.Y. [a.o.], 1988. B.B. Безуглов
|


