СПЛАЙСИНГ
(от англ.
splice-соединять, сращивать), удаление из молекулы РНК нитронов (участков РНК,
к-рые практически не несут генетич. информации) и соединение оставшихся участков,
несущих генетич. информацию (экзо-нов), в одну молекулу.
С.-один из этапов образования
функциональноактивных молекул РНК (процессинг РНК) из их предшественников, к-рый
осуществляется после завершения транскрипции
(синтез РНК на ДНК-матрице).
В результате удаления каждого интрона происходит разрыв двух фосфодиэфирных
связей с последующим образованием одной новой (см. Нуклеиновые кислоты
).
С. подвергаются предшественники
подавляющего большинства матричных РНК (пре-мРНК), а также нек-рых транспортных
и рибосомных РНК (соотв. пре-т РНК и пре-рРНК). С. характерен для РНК эукариот
(все организмы, за исключением бактерий и синезеленых водорослей); известны
также случаи С. РНК бактериофагов.
Механизмы С. у разл. классов
РНК различаются между собой. Для всех них характерна точность удаления интронов
и соединения экзонов. Специфичность удаления единств. интрона, если он имеется
в пре-тРНК, обеспечивается ее трехмерной структурой. Эндонуклеаза, ассоциированная
с ядерной мембраной, с участием др. ферментов расщепляет предшественник на участках
(сайтах) по краям интрона с образованием на концах экзонов 2,3-циклофосфатного
и 5-гидроксильного концов (рис. 1). Соединение этих концов осуществляется в
неск. стадий: у растений и дрожжей фосфорилирование 5-конца в месте разрыва
молекулы, превращение 2,3-циклофосфата в 2-фосфат и образование 3,5 -фосфодиэфирной
связи с участием остатка фосфорной к-ты из АТФ (левая часть рис.); у позвоночных
механизм С. пре-тРНК не включает фосфорилирование экзонов в месте разрыва (правая
часть рис.; на схеме указаны ферменты, катализирующие осн. этапы С.).
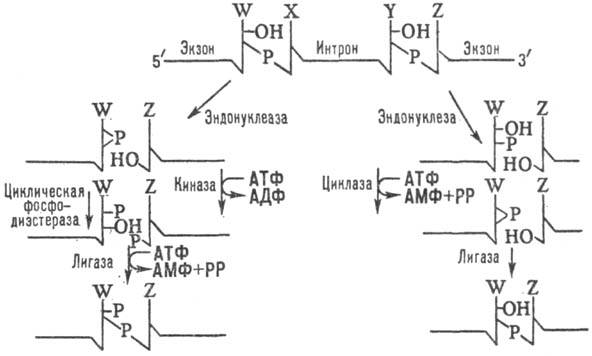
Рис. 1. Механизм сплайсинга
пре-тРНК; W, X, Y, Z-пуриновые или пири-мидиновые основания; АДФ,
АМФ, РР и P-соотв. аденозиндифосфат, аде-нозинмонофосфат, пирофосфорная
к-та и остаток фосфорной к-ты.
С. нек-рых пре-р РНК происходит
автокаталитически (аутосплайсинг, самосплайсинг). В этом случае катализатор
процесса - удаляемая интронная последовательность (рибо-зим). При этом С. осуществляется
в результате серии после-доват. р-ций, включающих превращение одного фосфоэфира
в другой без промежут. гидролиза фосфодиэфирных связей и использования энергии
извне. Р-ция происходит в присут. одновалентных катионов, Mg2+ и
гуанозинового кофактора (гуанозинового нуклеотида или самого гуанозина), к-рый
инициирует серию превращений - высвобождение интрона, соединение двух экзонов,
а также циклизацию интрона (при этом гуанозиновый кофактор регенерируется).
Самосплайсинг происходит
у пре-рРНК простейших (напр., у тетрахимоны) и ряда пре-мРНК митохондрий низших
грибов и нек-рых др. пре-РНК, у к-рых интроны содержат консервативные последовательности,
что обусловливает образование определенных вторичной и третичной структур.
Установлено участие при
удалении ряда интронов мито-хондриальных пре-мРНК у низших грибов особых белков-матюраз,
к-рые кодируются частично нитронами, частично экзонами. Роль матюраз, как и
нек-рых др. белков, сводится, по-видимому, к фиксации конформации интрона, необходимой
для осуществления им каталитич. ф-ции.
С. пре-мРНК, находящихся
в ядре, происходит в составе специфич. нуклеопротеидных частицах (сплайсомах).
Обычно С. подвергается кэпированная полиаденилированная линейная пре-мРНК. К.-л.
строгого порядка для удаления множественных интронов из пре-мРНК не наблюдается,
хстя удаление одних интронов может происходить быстрее, чем других. С. происходит
исключительно в ядре; несплай-сированная РНК остается в ядре и деградирует.
Однако если пре-РНК содержит интрон, к-рый может участвовать в альтернативном
пути С. (см. ниже), то она м. б. транспортирована в цитоплазму. С. ядерных пре-мРНК
происходит обычно по границам интронов, к-рые содержат на концах динуклеотиды
5-GU и AG-3 (A, G и U-соотв. остатки аденозина, гуанозина и уридина; правило
Шамбо-на). Известно только неск. исключений, когда вместо GU расположен динуклеотид
GC (С-остаток цитидина). Рядом с этими динуклеотидами расположены т. наз. консенсусные·
последовательности, к-рые имеют близкое строение у разл. пре-мРНК. Общая схема
С. ядерных пре-мРНК показана на рис.
2.
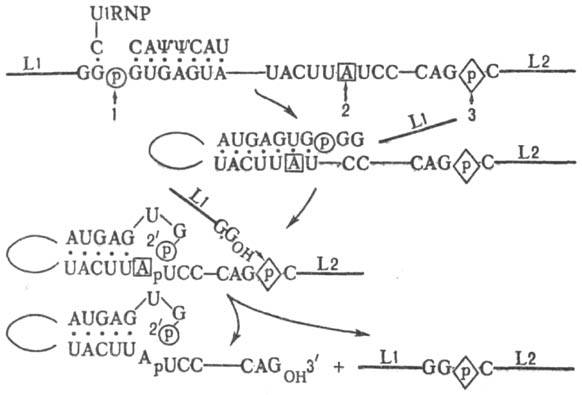
Рис. 2. Схема сплайсинга
пре-мРНК в ядрах клеток высших эукариот: 1 и 3-соотв. 5- и 3-концы интрона;
2-место разветвления; Y-остаток псевдоуридина (отсутствует атом N в положении
1 гетероцикла основания). Толстые линии-экзоны (L1 и L2), тонкие-интроны.
Для мн. пре-РНК известны
альтернативные пути С., дающие множественные формы зрелой РНК из транскрип-тов
одного гена. Это может иметь значение как один из механизмов регуляции экспрессии генов
, а также как ср-во увеличения кодирующей емкости генома (экспрессия
одного гена может
выражаться в синтезе разных мРНК). Известен также транс-С. (межмолекулярный
С.), при к-ром происходит соединение двух экзонов из разных молекул РНК.
Нарушение правильного С.
в результате мутаций, затрагивающих нуклеотидные последовательности ок. границ
интронов или экзонов, м. б. причиной возникновения наследственных болезней;
нарушенный С. у пре-мРНК арги-нинсукцинатсинтетазы приводит к цитрулйнемии,
пре-мРНК глобинов-к разл. типам талассемий, пре-мРНК иммуноглобулинов-к заболеваниям,
связанным с нарушением синтеза тяжелых цепей антител и др.
Лит.: Кавсан В.М.,
"Молекулярная биология", 1986, т. 20, в. 1, с. 5-20; там же, 1986.
в. 6, с. 1451-71; Padgett R. А. [а. о.], "Ann. Rev. Biochem.", 1986,
v. 55, p. 1119-50; Luhrmann R. [а.о.], Biochim. et Biophys. Acta",
1990, v. 1087, p. 265-92; Cech T. R., "Angew. Chemie", 1990, v.
29, № 7, p. 759-68; Altman S., там же, р. 749-58. В. М. Кавсан.
|


