ТРИКАРБОНОВЫХ КИСЛОТ ЦИКЛ
(цикл Кребса), цик-лич. последовательность ферментативных
р-ций (схема 1; назв. неионизир. форм к-т см. в ст. Обмен веществ
), в
к-рых осуществляются превращения ди- и трикарбоновых к-т, образующихся как промежут.
продукты в организме животных, в растениях и микробах.
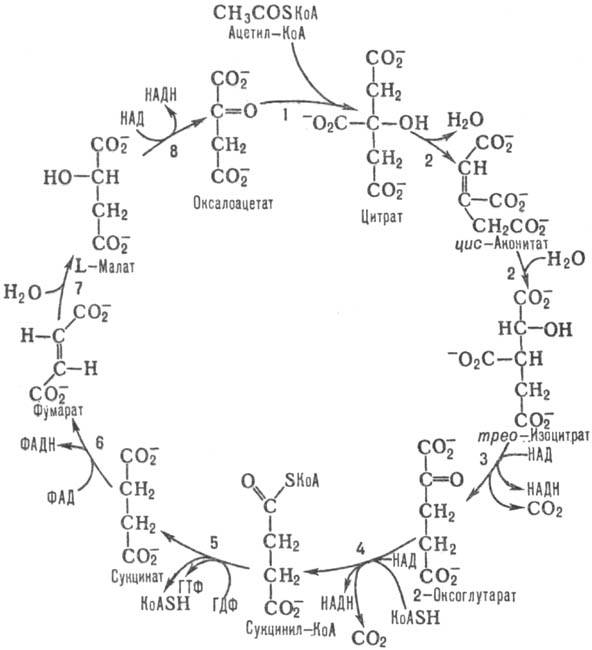
Схема 1. Цикл трикарбоновых
к-т.
Одновременно Т. к. ц.-метаболии,
путь окисления до СО2 и Н2О аминокислот, жирных к-т и
углеводов, к-рые вступают в этот цикл на разл. его стадиях (схема 2). Кроме
того, образующиеся ди- и трикарбоновые к-ты м. б. исходными субстратами в биосинтезе
мн. соед. (схема 3). Так, оксало-ацетат-субстрат в глюконеогепезе;
сукцинил-КоА - проме-жут. продукт в синтезе порфиринов, ацетил-КоА-в синтезе
жирных к-т, стероидов, ацетилхолина. Образующийся в цикле СО2 используется
в р-циях карбоксилирования в синтезе жирных к-т, орнитиновом цикле
и
др. Участие Т. к. ц. в биосинтезе и катаболизме мн. в-в обусловливает его важное
место в обмене в-в.
Т. к. ц. широко распространен
у всех аэробных организмов, у эукариот (все организмы, за исключением бактерий
и синезеленых водорослей) он осуществляется в митохондриях.
Суммарная р-ция Т. к. ц.
у животных имеет вид:
CH3C(O)SKoA
+ 3НАД + ФАД + ГДФ + F + Н2О : : 2СО2
+ 3НАДН + ФАДН + ГТФ + 2Н + KoASH
НАДН и НАД, ФАДН и ФАД-соотв.
восстановленные и окисленные формы кофермента никотинамидадениндинук-леотида
(см. Ниацин
) и кофермента флавинадениндинуклео-тида (см. Рибофлавин
);
ГДФ и ГТФ-соотв. гуанозинди-и гуанозинтрифосфат, Ф-неорг. фосфат, KoASH-кофер-мент
А.
НАДН и ФАДН, образующиеся
в цикле, окисляются в цепи переноса электронов (см. Дыхание
, Окислительное фосфорилирование
) с образованием АТФ, к-рый играет важную роль в
энергетич. обмене.
В р-ции 1 цикла, катализируемой
цитрат - оксалоацетат-лиазой, CH3C(O)SKoA стереоспецифично конденсируется
с карбонильной группой оксалоацетата с образованием цитрата и свободного KoASH.
Р-ция сопровождается значит. изменением своб. энергии (DG0
— 32,24 кДж/моль) и является практически необратимой. Активность митохонд-риального
фермента у дрожжей ингибируется АТФ.
Р-ция 2 цикла, катализируемая
аконитат-гидратазой,-изомеризация цитрата в изоцитрат путем последоват. дегидратации
- регидратации через промежут. образование цис
-аконитата. Р-ция обратима,
равновесие сдвинуто в сторону синтеза цитрата, однако в условиях непрерывного
функционирования цикла конечным продуктом р-ции является изоцитрат.
В р-ции 3, катализируемой
НАД- или НАДФ-зависимой изоцитратдегидрогеназой, происходит дегидрирование изо-цитрата
при атоме С-2 с одновременным декарбоксили-рованием и образованием 2-оксоглутарата
и СО2. Бактерии содержат НАДФ-зависимую изоцитратдегидрогеназу, активность
к-рой регулируется хим. модификацией - фосфо-рилированием (инактивация) и дефосфорилированием
(активация) при участии бифункцион. фермента изоцитратдегид-рогеназа-киназа
(фосфатаза), играющего существ. роль в переключении обмена с Т. к. ц. на анаплеротич.
("возмещающий" образование промежут. продуктов Т. к. ц.) глиокси-латный цикл. Эукариоты содержат обе формы изоцитрат-дегидрогеназы. Активность НАДФ-зависимого
фермента, локализованного в митохондриальном матриксе и цитозоле, контролируется
продуктами р-ции. Активность НАД-зави-симой изоцитратдегидрогеназы, локализованной
исключительно в митохондриальном матриксе, активируется у грибов аденозинмонофосфатом
(АМФ) и цитратом, у животных - аденозиндифосфатом (АДФ), цитратом и ионами Са2+.
Р-ция 4 катализируется
мультиферментным 2-оксоглута-ратдегидрогеназным комплексом, состоящим из трех
осн. ферментов: 2-оксоглутаратдегидрогеназы, дигидролипоил-сукцинилтрансферазы
и дигидролипоилдегидрогеназы.
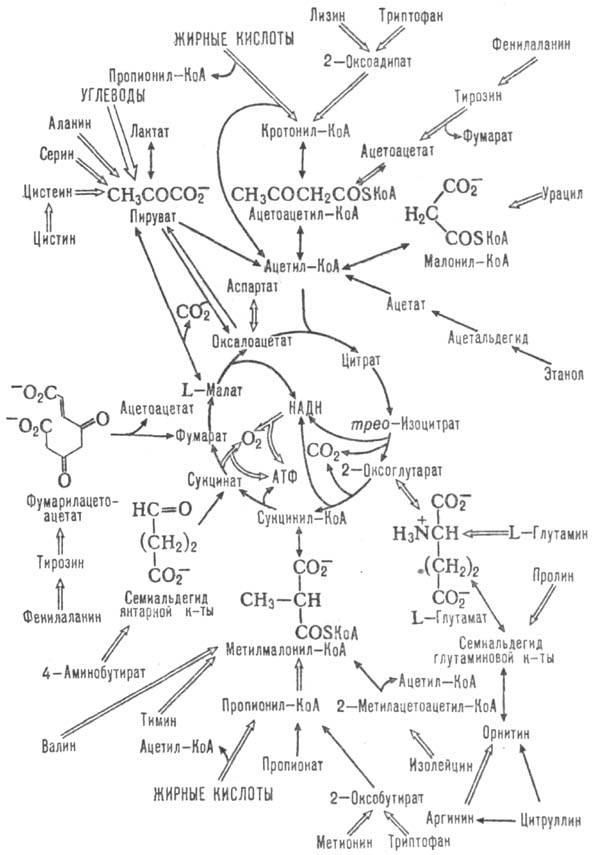
Схема 2. Цикл трикарбоновых
к-т и катаболич. р-ции. Двойными стрелками отмечены многостадийные р-ции; семиальдегиды
к-т w-оксокислоты.
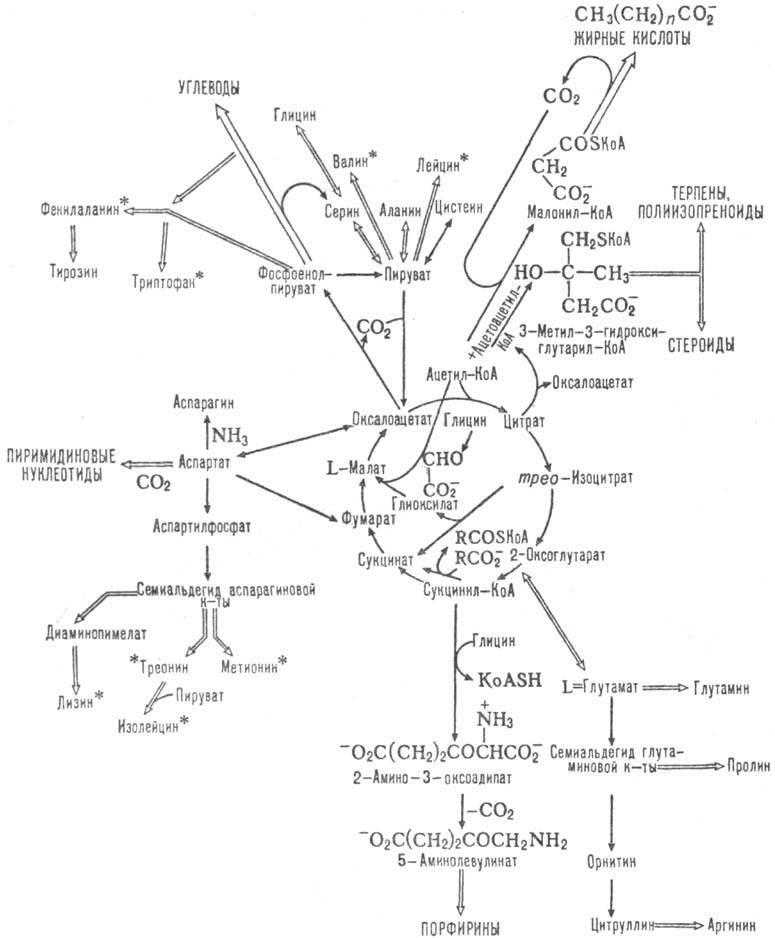
Схема 3. Биосинтетич. и
др. р-ции, сопутствующие циклу трикарбоновых к-т; аминокислоты, отмеченные звездочкой,
в организме высших животных несинтезирую гея.
Сукцинил-КоА в р-ции 5,
катализируемой сукцинил-КоА -синтетазой, подвергается распаду, в результате
к-рого энергия тиоэфирной связи сукцинил-КоА запасается в виде синтезир. нуклеозидтрифосфата
(у бактерий, грибов, растений-АТФ, у животных - ГТФ).
В р-ции 6, катализируемой
сукцинатдегидрогеназой
, происходит превращение сукцината в фумарат. Фермент
входит в состав более сложного сукцинатдегидрогеназного комплекса (комплекса
II) дыхат. цепи, поставляя восстановит. эквиваленты, образующиеся в р-ции, в
дыхат. цепь.
Фумарат-гидратаза, катализирующая
р-цию 7, осуществляет гидратирование фумарата с образованием L-малата. Активность
фермента ингибируется АТФ.
Последняя р-ция цикла,
8, катализируется L-малатдегид-рогеназой; L-малат при этом превращ. в оксалоацетат,
к-рый может взаимод. с новой молекулой ацетил-КоА. Р-ция обратима, равновесие
сдвинуто в сторону образования L-малата (DG0 +29,73
кДж/моль), но в условиях функционирования цикла конечным продуктом р-ции является
оксалоацетат.
Предполагают, что катализируемые
индивидуальными ферментами р-ции осуществляются благодаря действию надмолекулярного
"сверхкомплекса", т. наз. метаболона. Преимущества такой организации
ферментов очевидны -при этом не происходит диффузии кофакторов и субстратов,
что способствует более эффективной работе цикла.
Наличие в Т. к. ц. 4 восстановит.
р-ций (3, 4, 6 и 8), в результате к-рых на 1 молекулу CH3C(O)SKoA
синтезируются 3 молекулы НАДН и 1 молекула ФАДН, определяет необходимые
условия для его функционирования. Непрерывная работа цикла требует реокисления
НАДН и ФАДН, к-рое в аэробных условиях, как правило, осуществляется через совокупность
переносчиков электронов, составляющих дыхат. цепь, и сопровождается запасанием
значит. кол-ва энергии. У животных это 11 молекул АТФ на 1 молекулу окисленного
ацетил-КоА. У грибов, растений и особенно бактерий число молекул АТФ, образующихся
при окислении НАДН и ФАДН, м.б. меньше вследствие разветвления дыхат. цепи.
В анаэробных условиях вместо
Т.к.ц. функционируют его окислит. ветвь до 2-оксоглутарата (р-ции 1 :
2 : 3) и восстановительная - от оксалоацетата до сукцината (р-ции 8 :
7 : 6). При этом не происходит запасания большого кол-ва энергии и ф-ция
цикла целиком определяется доставкой в-в для синтеза клеточного материала.
При переходе организма
от покоя к активному состоянию возникает потребность в ускоренной мобилизации
энергии (обменных процессов). У животных это достигается, в частности, шунтированием
наиб. медленных р-ций Т. к. ц. (р-ции 1-3) и преимуществ. окислением сукцината.
При этом исходный субстрат укороченного Т. к. ц. (2-оксоглутарат) образуется
в результате быстрой р-ции переаминирования:
Глутамат + Оксалоацетат
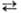 2-Оксоглутарат
+ Аспартат
2-Оксоглутарат
+ Аспартат
Связь Т. к. ц. с глиоксилатным
циклом осуществляется благодаря синтезу в последнем сукцината, к-рый в Т. к.
ц. окисляется до оксалоацетата и служит, т. обр., поставщиком молекул с 4 атомами
С в оба цикла на начальных стадиях их функционирования. Благодаря этому возможно
функционирование этих циклов, когда в организме избыток соед. с 2 атомами С,
напр. при выращивании бактерий на средах с СН3СООН и С2Н5ОН,
а также при прорастании семян масличных растений, во время к-рого усиленно образуется
ацетил-КоА.
Др. модификация Т. к. ц.
(т. наз. 4-аминобутиратный шунт) - превращение 2-оксоглутарата в сукцинат, через
глу-таминовую к-ту, 4-аминобутират и янтарный семиальдегид (3-формилпропионовая
к-та). Эта модификация имеет большое значение для ткани мозга, в к-рой ок. 10%
глюкозы распадается по этому пути.
Тесное сопряжение р-ций
Т. к. ц. с дыхат. цепью, особенно в митохондриях животных, а также особенности
регуляции активности индивидуальных ферментов цикла (для большинства из них
АТФ является ингибитором) предопределяют снижение активности цикла в условиях
генерирования высокого фосфорильного потенциала (отношения АТФ/АДФ) в клетке,
и наоборот - активацию цикла при пониж. фосфорильном потенциале. У большинства
растений, бактерий и мн. видов грибов тесное сопряжение цикла с дыхат. цепью
преодолевается развитием альтернативных несопряженных путей окисления, позволяющих
поддерживать дыхат. активность и активность Т. к. ц. на высоком уровне даже
в условиях высокого фосфорильного потенциала. Т. к. ц. открыт в 1937 X. Кребсом
и У. Джонсоном.
Лит.: Страйер Л.,
Биохимия, пер. с англ., т. 2, М., 1985, с. 49-68; Ленинджер А., Основы биохимии,
пер. с англ., т. 2, М., 1985, с. 477-507; Кондрашова М.Н., "Биохимия",
1991, т. 56, в. 3, с. 388-404; Krebs H. А., Kornberg H. L., Energy
transformations in living matter, В., 1957; Krebs H. А., "Persp.
Biol. Med.", 1970, v. 14, p. 154-70; Srere P. A., "Ann. Rev. Biochem.",
1987, v. 56, p. 89-124. P. А. Звягильская.
|


